Bovine respiratory syncytial virus (BRSV) is a major cause of bronchiolitis and pneumonia in cattle and causes yearly outbreaks with high morbidity in Europe. Commercial vaccines against this virus needs improvement of efficacy, especially in calves with BRSV-specific maternally derived antibodies (MDA). We previously reported that an experimental BRSV-ISCOM vaccine, but not a commercial vaccine, induced strong clinical and virological protection in calves with MDA, immunized at 7–15 weeks of age. The aim of the present study was to characterize the immune responses, as well as to investigate the efficacy and safety in younger animals, representing the target population for vaccination. Four groups of five 3–8 week old calves with variable levels of BRSV-specific MDA were immunized s.c. twice at a 3 weeks interval with (i) BRSV immunostimulating complexes (BRSV-ISCOMs), (ii) BRSV-protein, (iii) adjuvant, or (iv) PBS. All calves were challenged with virulent BRSV by aerosol 2 weeks later and euthanized on day 6 after infection. The cellular and humoral responses were monitored as well as the clinical signs, the viral excretion and the pathology following challenge. Despite presence of MDA at the time of the immunization, only a minimum of clinical signs were observed in the BRSV-ISCOM group after challenge. In contrast, in all control groups, clinical signs of disease were observed in most of the animals (respiratory rates up to 76 min−1 and rectal temperatures up to 41 °C). The clinical protection was associated to a highly significant reduction of virus replication in the upper and lower respiratory tract of calves, rapid systemic and local antibody responses and T helper cell responses dominated by IFNγ production. Animals that did not shed virus detectable by PCR or cell culture following challenge possessed particularly high levels of pulmonary IgA. The protective immunological responses to BRSV proteins and the ability to overcome the inhibiting effect of MDA were dependent on ISCOM borne antigen presentation.
Excerpt from my doctoral defense presentation
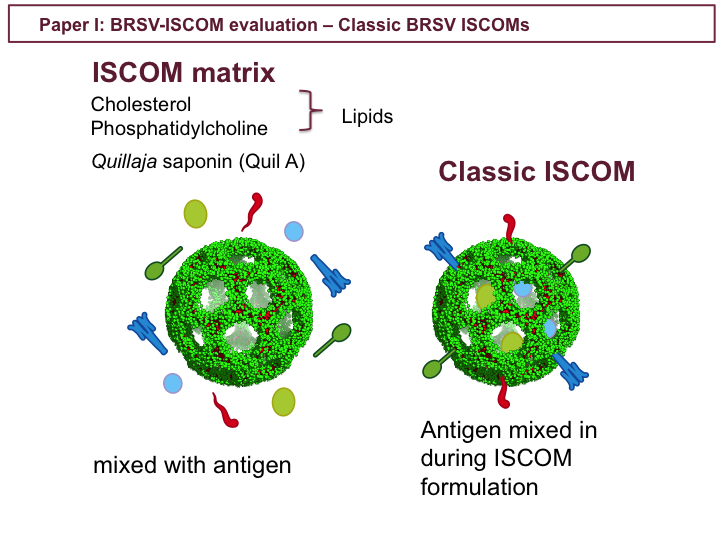
In paper I classic BRSV-ISCOMs were evaluated. ISCOM matrices are spherical structures formed when certain lipids and saponin are mixed under specific conditions. ISCOM matrices, such as the AbISCO-300 used in study IV, are commercially available. ISCOMs have been shown to boost immune responses to co-delivered antigens, and induce humoral and both Th1 and Th2 type T-cell responses. If antigens are included in the mix when the ISCOMs are formed, some antigens may be more integrated into the ISCOMs. This is referred to as classic ISCOMs, and may enhance the immunogenicity of the formulation. (ISCOM matrix image from: Pedersen et al. Biophysical Journal 2012 102, 2372-2380)
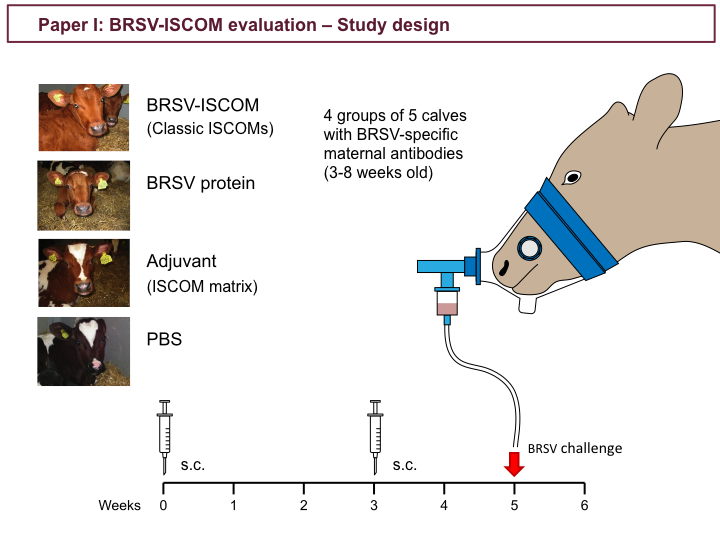
For paper I, classic BRSV-ISCOMs were formed using purified antigens from lysate of BRSV-infected cells. To evaluate the the safety, immunogenicity and protective efficacy of these BRSV-ISCOMs, 4 groups of 5 calves with specific maternal antibodies were immunized subcutaneously twice at an interval of 3 weeks with either: BRSV-ISCOMs, BRSV protein alone, ISCOM matrix adjuvant alone, or Phosphate-buffered saline solution (PBS). Five weeks after first immunization, all calves were challenged by aerosol inhalation of BRSV, and monitored for 6 days.
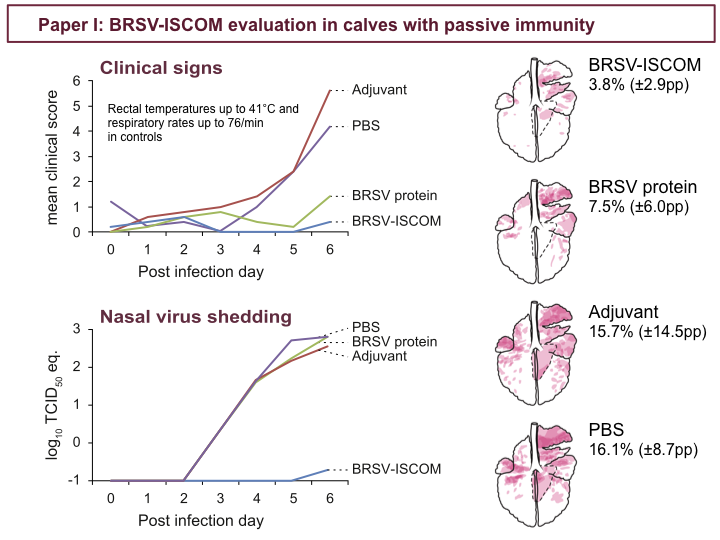
Following challenge, calves immunized with Adjuvant or PBS demonstrated moderate to severe clinical signs, as seen in the upper chart showing clinical scores after challenge. All calves except calves immunized with BRSV-ISCOMs shed high amounts of virus in nasal secretion, detected by quantitative PCR, shown in the lower chart. In contrast, no or minimal levels of virus RNA was detected in BRSV-ISCOM-immunized calves. BRSV-ISCOM-immunized calves were also significantly protected from lung pathology, as seen in these per-group lung charts of superimposed individual lung charts. Overall, it was obvious that BRSV-ISCOMs induced a high level of protection. To understand what mediated this protection, we investigated both humoral and cellular immune parameters.
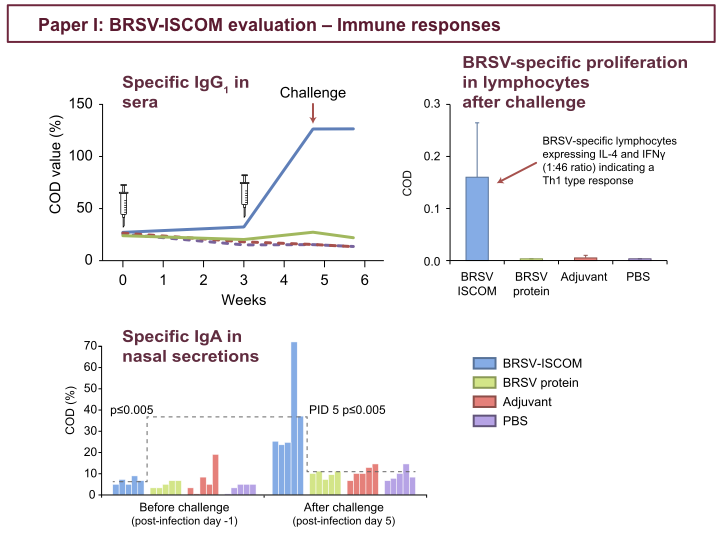
In the upper left figure, you see increasing levels of BRSV-specific IgG1 in sera of BRSV-ISCOM-vaccinated calves, already after first vaccination, which accelerate sharply after boost. In contrast, control calves have declining levels of specific MDA throughout the experiment. Strikingly, BRSV-ISCOM-immunized calved demonstrated a rapid increase of BRSV-specific IgA antibodies in nasal secretions after challenge, as shown in the lower figure, but also in bronchoalveolar lavage (BAL) at post-infection day (PID) 6. The figure to the right illustrates that BRSV-ISCOMs also primed BRSV-specific T-cells in vaccinated calves. These lymphocytes were also shown to express interleukin 4, or IL-4, and interferon gamma, and the predominance of IFN gamma suggested a Th1-biased T-cell response.
In conclusion for paper I, classic BRSV-ISCOMs overcame the suppressive effect of specific maternal antibodies and induced a high level of protection in young calves, and this protection was associated with rapid BRSV-specific mucosal IgA responses in the upper and lower airways, and a Th1-oriented T cell response. These finding were published in Vaccine in 2011.
As mentioned, the BRSV-ISCOMs were produced using purified antigen from lysate of BRSV-infected cells, and in this purification process, only one fraction of the total antigen is used in the final vaccine. The exceptional protection induced by BRSV-ISCOMs leads to the question: What’s in the BRSV-ISCOMs? This question was addressed in Paper II.