Vaccine Safety and Efficacy Evaluation of a Recombinant Bovine Respiratory Syncytial Virus (BRSV) with Deletion of the SH Gene and Subunit Vaccines Based On Recombinant Human RSV Proteins: N-nanorings, P and M2-1, in Calves with Maternal Antibodies
Krister Blodörn, Sara Hägglund, Jenna Fix, Catherine Dubuquoy, Boby Makabi-Panzu, Michelle Thom, Per Karlsson, Jean-Louis Roque, Erika Karlstam, John Pringle, Jean-François Eléouët, Sabine Riffault, Geraldine Taylor, Jean François Valarcher PLoS ONE 06/2014; 9(6):e100392. DOI:10.1371/journal.pone.0100392
Abstract
The development of safe and effective vaccines against both bovine and human respiratory syncytial viruses (BRSV, HRSV) to be used in the presence of RSV-specific maternally-derived antibodies (MDA) remains a high priority in human and veterinary medicine. Herein, we present safety and efficacy results from a virulent BRSV challenge of calves with MDA, which were immunized with one of three vaccine candidates that allow serological differentiation of infected from vaccinated animals (DIVA): an SH gene-deleted recombinant BRSV (ΔSHrBRSV), and two subunit (SU) formulations based on HRSV-P, -M2-1, and -N recombinant proteins displaying BRSV-F and -G epitopes, adjuvanted by either oil emulsion (Montanide ISA71VG, SUMont) or immunostimulating complex matrices (AbISCO-300, SUAbis). Whereas all control animals developed severe respiratory disease and shed high levels of virus following BRSV challenge, ΔSHrBRSV-immunized calves demonstrated almost complete clinical and virological protection five weeks after a single intranasal vaccination. Although mucosal vaccination with ΔSHrBRSV failed to induce a detectable immunological response, there was a rapid and strong anamnestic mucosal BRSV-specific IgA, virus neutralizing antibody and local T cell response following challenge with virulent BRSV. Calves immunized twice intramuscularly, three weeks apart with SUMont were also well protected two weeks after boost. The protection was not as pronounced as that in ΔSHrBRSV-immunized animals, but superior to those immunized twice subcutaneously three weeks apart with SUAbis. Antibody responses induced by the subunit vaccines were non-neutralizing and not directed against BRSV F or G proteins. When formulated as SUMont but not as SUAbis, the HRSV N, P and M2-1 proteins induced strong systemic cross-protective cell-mediated immune responses detectable already after priming. ΔSHrBRSV and SUMont are two promising DIVA-compatible vaccines, apparently inducing protection by different immune responses that were influenced by vaccine-composition, immunization route and regimen.
Excerpt from my doctoral defense presentation
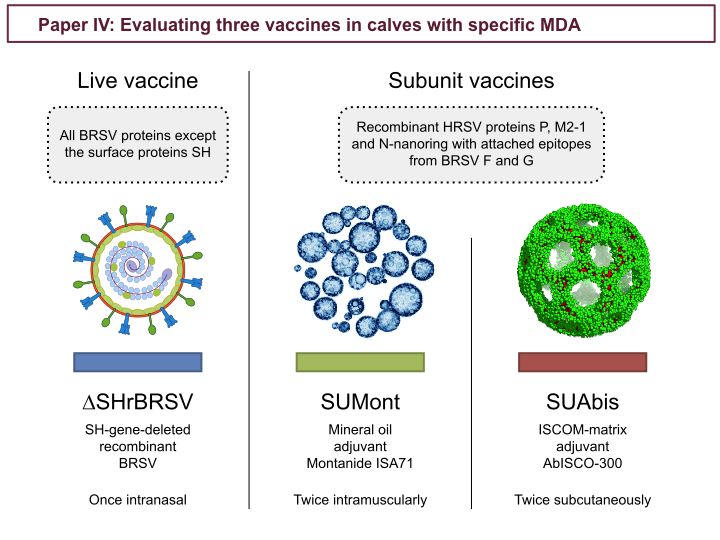
In study IV three vaccines candidates were evaluated: one recombinant live BRSV with the SH-gene deleted (∆SHrBRSV), and two subunit vaccines, with the same protein subunit composition, but formulated by either with: a Montanide mineral oil emulsion (SUMont) or AbISCO-300 ISCOM-matrices (SUAbis). The subunit composition was designed using information gained from characterization of the BRSV-ISCOMs in study II, but was restricted by available constructs. Both subunit vaccines contained recombinant HRSV proteins P and M2-1. But, because we were unable to produce a full-length recombinant F protein, we instead used recombinant nanorings of the HRSV N protein, with grafted epitopes from BRSV proteins F and G (see introduction about BRSV proteins). (ISCOM matrix image from: Pedersen et al. Biophysical Journal 2012 102, 2372-2380)
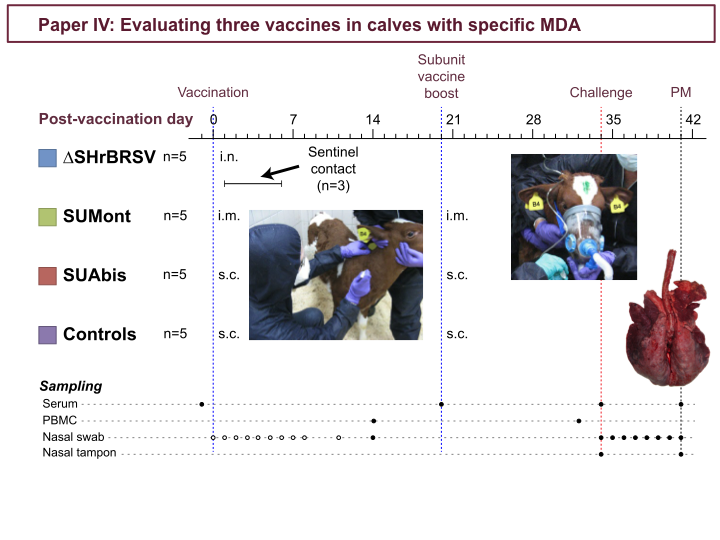
In study 4, three vaccines were evaluated. Four groups of 5 calves with specific MDA were vaccinated 5 weeks before challenge: the ∆SH intranasally, and the others parenterally, as indicated in the timeline. In addition, subunit-vaccination was boosted three weeks later. Also, 3 seronegative calves, were co-housed with the ∆SH-calves, and acted as sentinels for transmission of the vaccine virus. But there was no indication of transmission, as none of the sentinels seroconverted, showed any clinical signs, or shed any detectable virus. Five weeks after first vaccination, all calves were challenged with BRSV by aerosol inhalation (as described in detail in paper III) and then monitored for 7 days.
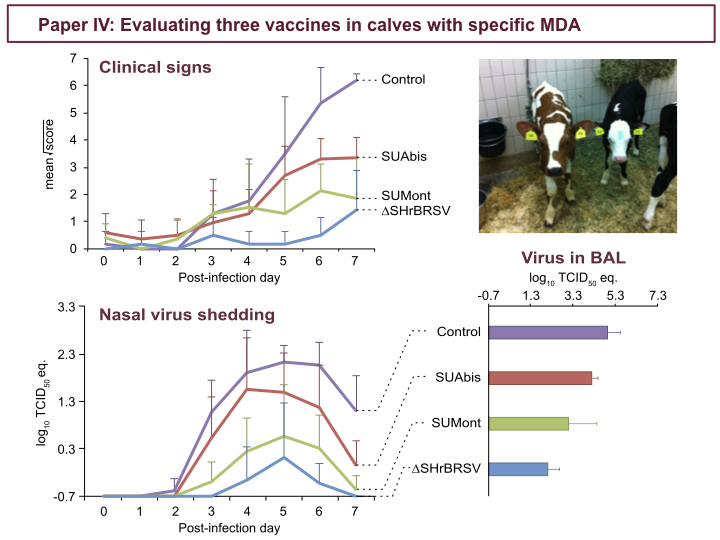
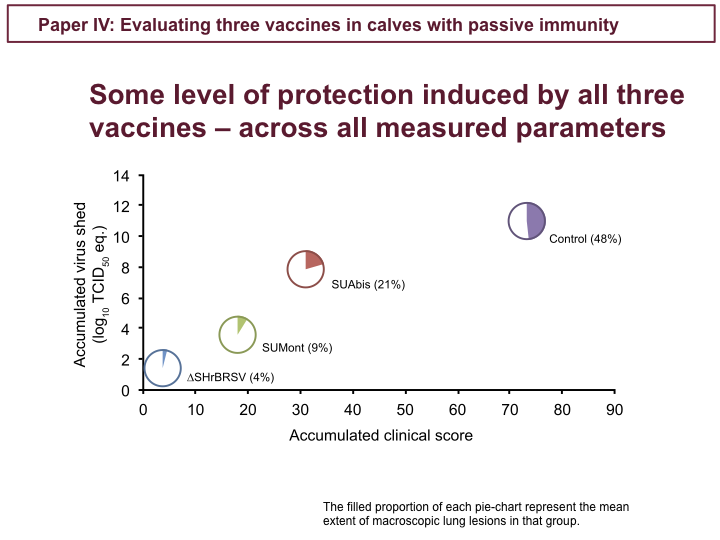
These figures show clinical scores and virus detected by PCR in nasal secretion, during the 7 days following challenge, as well as virus in the lungs on PID 7. In contrast to control calves, in purple, all the vaccinated calves were afforded some level of clinical protection. The ∆SH-vaccinated calves, in blue, were almost completely clinically protected, while the SUMont-vaccinated calves, in green, were well protected, and the SUAbis-vaccinated calves, in red, were least protected. Similarly for virus shed, while controls, in purple, shed high amounts of virus in nasal secretion, we see the same pattern of virological protection among vaccinated groups: the ∆SH, in blue, very well protected, the SUMont, in green, well protected, and the SUAbis, in red, the least virologically protected. Again, we see the same pattern among groups, when looking at virus detected by PCR in the BAL, in the lower right.
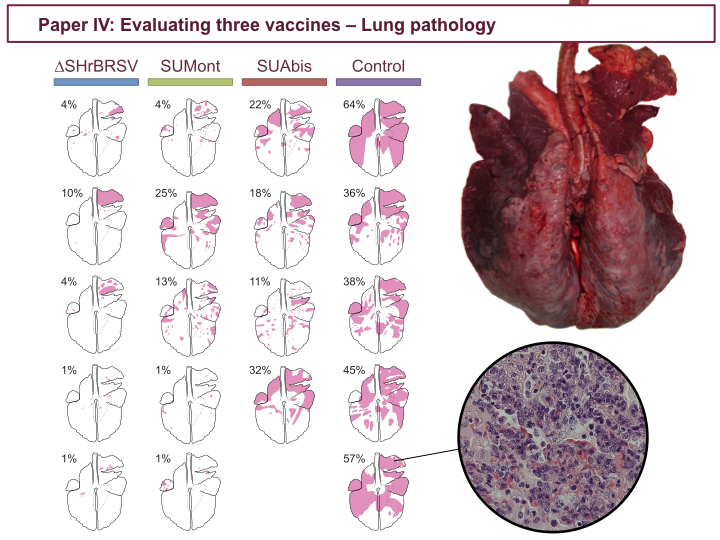
The data presented in the previous figures also correlates with the degree of consolidative lesions in the lungs at post-mortem, indicated in pink in these lung charts. Whereas controls, to the right, had massive lesions involving up to 64% of the lung tissue, calves vaccinated once intranasally with ∆SH, to the left, had only small and scattered lesions. Again, SUMont-vaccinated calves appeared well protected, and SUAbis-vaccinated animals were least protected. The histopathological severity of lesions correlated with the extent of densification.
Across clinical, pathological and virological parameters control calves were severely affected, whereas the ∆SH-vaccinated calves were very well protected, SUMont calves were well protected, and SUAbis calves were the least protected. So, how do these distinct levels of protection correlate with immunological responses to vaccination?
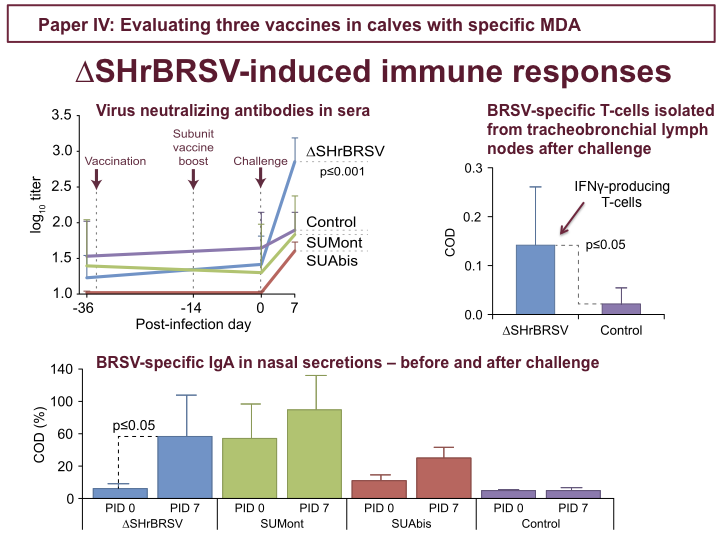
For ∆SHrBRSV, there was no indication of any immune responses in the 5 weeks following intranasal vaccination. However, following challenge, ∆SH-calves demonstrated rapidly increasing titers of neutralizing antibodies in sera, as shown in the upper figure, and BRSV-specific IgA in nasal secretion, as seen in the lower figure, but also in BAL. In addition, ∆SH-calves demonstrated priming of IFNγ-producing BRSV-specific T-cells in tracheobronchial lymph nodes analyzed after challenge. The subunit vaccines on the other hand did not induce neutralizing antibodies, and although SUMont induced BRSV-specific IgA in nasal secretions, detectable already before challenge, these were likely also non-neutralizing.
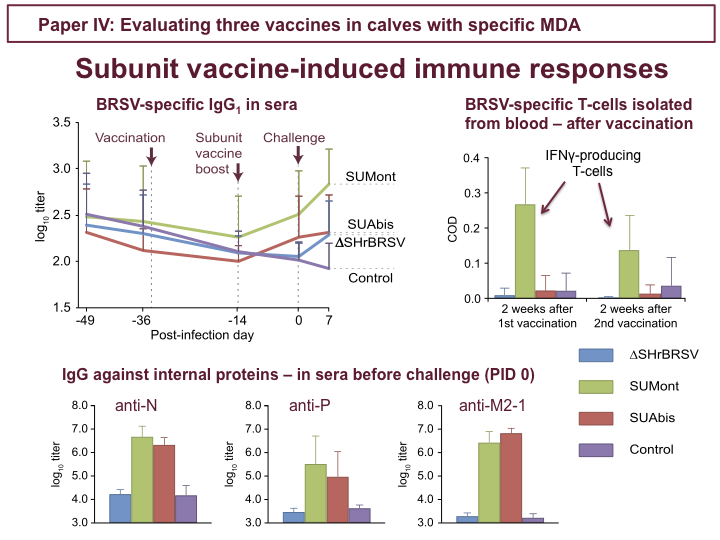
In fact, although both subunit vaccines induced humoral responses detected by BRSV-specific IgG1 ELISA, as shown in the upper figure, these were directed against the internal proteins included in both vaccines: N, P and M2-1, as shown in the lower figures, showing titers just before challenge. Results from F and G protein-specific ELISAs suggested that the BRSV F- and G-epitopes included in the subunit vaccines did not contribute to protection. Unfortunately, tracheobronchial lymph nodes from subunit-vaccinated calves could not be processed at post-mortem, but when lymphocytes isolated from blood, before challenge, were restimulated with BRSV SUMont-vaccinated calves demonstrated strong priming of BRSV-specific IFNγ-producing T-cells, 2 weeks after both first and second vaccination, as indicated in the figure to the right. The SUAbis vaccine induced humoral responses similar to that induced by SUMont, but lower amplitude, and no detectable cellular response.
To conclude study IV, we identified two good candidate vaccines: The ∆SH recombinant BRSV, where we saw nothing before challenge, but which primed for strong anamnestic mucosal and neutralizing serum antibodies and local BRSV-specific T-cells, and which afforded almost complete protection after a single intranasal vaccination; and SUMont, which induced both T-cell and humoral immune responses directed against HRSV proteins N, P and M2-1, and a good level of protection.
I presented these findings at the RSV Vaccines for the World conference in Portugal in 2013, and they were later published in PLoS ONE.